
Функция жгутиков в бактериальной клетке
Содержание:
Читайте также:
- Жгутики
- Жгутики у бактерий. Размеры, форма, взаиморасположение. Механизм движения жгутиков. Ворсинки (фимбрии) и их функции.
Строение.Около половины известных видов бактерий на поверхности имеют органы движения — волнообразно изогнутые жгутики. Масса жгутиков составляет до 2 % сухой массы бактерии. Длина жгутика больше длины тела микроорганизма и составляет 3–12 мкм; толщина жгутика 0,02 мкм, причем полярные жгутики более толстые, чем перитрихиальные.
Жгутики состоят из белка флагеллина (лат. flagella — жгутик), который по своей структуре относится к сократительным белкам типа миозина. В составе жгутика имеется либо одна гомогенная белковая нить, либо 2–3 нити, плотно свернутые в косу. Нить жгутика — жесткая спираль, закрученная против часовой стрелки; шаг спирали специфичен для каждого вида бактерий.
Число, размеры и расположение жгутиков являются признаками, постоянными для определенного вида, и учитываются при систематике. Однако у некоторых бактерий могут образовываться жгутики разных типов. Кроме того, наличие жгутиков зависит от условий внешней среды: на твердых средах при длительном культивировании бактерии могут утратить жгутики, а на жидких — вновь приобрести. Количество и расположение жгутиков у одного и того же вида может определяться стадией жизненного цикла. Следовательно, не стоит переоценивать таксономическое значение этого признака.
Классификация бактерий по числу и расположению жгутиков:
1. Атрихи— жгутики отсутствуют.
2. Монотрихи — один жгутик, расположенный на одном из полюсов клетки (род Vibrio) — монополярное монотрихальное расположение жгутиков, самые подвижные бактерии.
3. Политрихи — много жгутиков:
– лофотрихи— пучок жгутиков на одном полюсе клетки(роды Pseudomonas, Burkholderia)—монополярное политрихальное расположение жгутиков;
– амфитрихи — на каждом полюсе клетки расположено по пучку жгутику (род Spirillum) — биполярное политрихальное расположение жгутиков;
– перитрихи — жгутики расположены без определенного порядка по всей поверхности клетки (сем. Enterobacteriaceae (роды Escherichia, Proteus), сем. Bacillaceae, сем. Clostidiaceae), число жгутиков от 6 до 1000 на клетку в зависимости от вида бактерий (рис. 7).

Рис.7. Варианты расположения жгутиков у бактерий:
1 — монотрих, 2 — лофотрих;
3 — амфитрих; 4 — перитрих.
– При электронной микроскопии обнаружено, что жгутик состоит из трех частей:спиральной нити, крюка и базального тела (рис. 8).
Основную часть жгутика составляет длинная спиральная нить (фибрилла) — жесткий полый цилиндр диаметром около 120 нм, состоящий из белка флагеллина. По длине нити белковые молекулы образуют 11 рядов и уложены в виде спирали. В процессе роста нити белковые молекулы, синтезированные внутри клетки, проходят через полость цилиндра и пристраиваются в спираль на ее конце. На конце жгутика имеется белковая шапочка (крышечка), закрывающая отверстие цилиндра и препятствующая выходу молекул белка в окружающую среду. Длина нити жгутика может достигать нескольких микрометров. У некоторых видов бактерий жгутик снаружи дополнительно покрыт чехлом. У поверхности КС спиральная нить переходит в утолщенную изогнутую структуру — крюк.
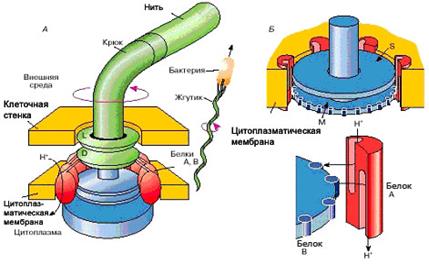
Рис. 8. Схема строения жгутика
2. Крюк (толщиной 20–45 нм) вблизи поверхности клетки — относительно короткий цилиндр, состоит из белка, отличающегося от флагеллина, и служит для обеспечения гибкого соединения нити с базальным телом.
3. Базальное тело находится в основании жгутика и обеспечивает его вращение. Базальное тело содержит 9–12 различных белков и состоит из двух или четырех дисков (колец), нанизанных на стержень, являющийся продолжением крюка. Эти кольца вмонтированы в ЦПМ и КС. Два внутренних кольца (M и S) — обязательные составные части базального тела. M-кольцо локализовано в ЦПМ, S-кольцо располагается в периплазматическом пространстве грамотрицательных или в пептидогликановом мешке грамположительных бактерий. Два наружных кольца (D и L) необязательны для движения, так как имеются только у грамотрицательных бактерий, локализованы соответственно в пептидогликановом слое и в наружной мембране КС. Кольца S, D и L неподвижны и служат для фиксации жгутика в КС. Вращение жгутика определяется вращением M–кольца, встроенного в ЦПМ клетки. Таким образом, особенности строения базального тела жгутика определяются строением КС.
Функционально базальное тело представляет собой электромотор, работающий на протонах. М–кольцо базального тела (вращающийся ротор), окружено мембранными белками, имеющими отрицательные заряды (статор мотора). Бактериальная клетка обладает эффективным механизмом, позволяющим превращать электрохимическую энергию в механическую. Поэтому на работу жгутика бактерия тратит около 0,1 % всей расходуемой ею энергии. При работе жгутика используется протондвижущая сила, которая обеспечивается разностью концентраций протонов на внешней и внутренней сторонах мембраны (на внешней стороне их больше) и наличием более отрицательного заряда на внутренней стороне мембраны. Протондвижущая сила заставляет протоны проходить через базальное тело внутрь клетки, при этом они задерживаются на определенных участках ротора, придавая им положительный заряд, затем протоны уходят внутрь клетки. Заряженные участки расположены таким образом, что возникает сила притяжения между заряженными участками ротора и статора, М-кольцо начинает вращаться со скоростью около 300 об/c. Механизм вращения: зарядка–перезарядка группы COOH в аминокислотах. Для полного оборота кольца через базальное тело должно пройти 500–1000 протонов. Вращение М-кольца через жестко связанную с ним ось и крюк передается нити жгутика, которая функционирует как пропеллер или корабельный винт. Бактерия плывет до тех пор, пока работает винт, вклад инерции исключительно мал.
Кроме того, бактерии, даже мертвые, находящиеся в водной среде, перемещаются в результате броуновского движения. Бактериальная клетка все время подвержена ударам окружающих молекул, находящихся в тепловом движении. Удары, наносимые с разных сторон, бросают бактерию из стороны в сторону.
Тип движения жгутиков — вращательный. Существуют два вида движения: прямолинейное и кувыркание (периодические случайные изменения направления движения). Когда жгутики вращаются против хода часовой стрелки (около 1 секунды), с частотой 40–60 об./с (близко к скорости среднего электромотора), их нити сплетаются в единый жгут (рис. 9а). Вращение жгутиков передается клетке. Так как клетка намного массивнее жгутика, она начинает двигаться по прямой в противоположном направлении, со скоростью в 3 раза меньшей, чем скорость движения жгутика.
Так обеспечивается поступательное движение клетки, скорость которого в жидкой среде для разных видов бактерий составляет 20–200 мкм/с (это соответствует примерно 300–3000 длин тела в минуту) и более медленное перемещение по поверхности твердых сред.
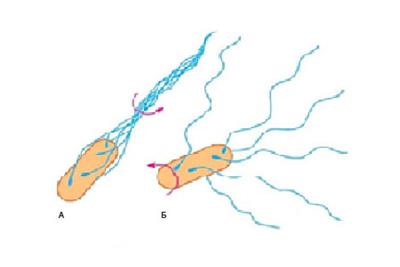 |
Рис. 9. Расположение жгутиков на клетке кишечной палочки при их вращении: А — против часовой стрелки, Б — по часовой стрелке |
Плыть целенаправленно в одном направлении бактерия может не более 3 с, затем удары окружающих молекул разворачивают ее в случайном направлении. Поэтому бактерии выработали механизм спонтанного изменения направления движения — переключение жгутикового мотора. Когда он начинает вращаться по ходу часовой стрелки (около 0,1 с), бактерия останавливается и переворачивается (совершает «кувырок») в случайном направлении. При этом жгутики разлетаются в разные стороны (рис. 9б). У амфитрихов при движении один пучок жгутиков вывернут наизнанку (подобно вывернутому ветром зонту). Затем мотор снова переходит к вращению против часовой стрелки, и бактерия опять плывет по прямой, но уже в другом, случайном направлении.
Жгутики могут также изменять направление движения в ответ на внешний стимул. Если бактерия перемещается в сторону оптимальной концентрации аттрактанта, жгутики проталкивают клетку через среду, ее прямолинейное движение становится более длительным, а частота кувырканий более низкой, что позволяет ей в конечном итоге перемещаться в нужном направлении.
Известны случаи существования бездействующих (парализованных) жгутиков. Для движения жгутиковых бактерий необходима интактность (неповрежденность) КС. Обработка клеток лизоцимом, приводящая к удалению пептидогликанового слоя КС, вызывает потерю способности бактерий к движению, хотя жгутики остаются при этом неповрежденными.
Таксисы бактерий. Пока окружающая среда остается неизменной, бактерии плавают беспорядочно. Однако совершенно однородной окружающая среда бывает редко. Если среда неоднородна, бактерии проявляют элементарные поведенческие реакции: активно перемещаются в направлении, определяемом теми или иными внешними факторами. Такие генетически детерминированные целенаправленные перемещения бактерий называют таксисами. В зависимости от фактора различают хемотаксис (частный случай — аэротаксис), фототаксис, магнитотаксис, термотаксис и вискозитаксис.
Хемотаксис — движение в определенном направлении относительно источника химического вещества. Химические вещества делят на две группы: инертные и вызывающие таксисы — хемоэффекторы. Среди хемоэффекторов есть вещества, привлекающие бактерий, — аттрактанты (сахара, аминокислоты, витамины, нуклеотиды), и вещества, их отпугивающие, — репелленты (некоторые аминокислоты, спирты, фенолы, неорганические ионы). Аттрактантом для аэробных и репеллентом для анаэробных прокариот является молекулярный кислород. Аттрактанты часто представлены пищевыми субстратами, хотя не все вещества, необходимые для организма, выступают в качестве аттрактантов. Также не все ядовитые вещества служат репеллентами и не все репелленты вредны. Таким образом, бактерии способны реагировать не на любые соединения, а только на определенные и различные для разных бактерий.
В поверхностных структурах бактериальной клетки есть специальные белковые молекулы — рецепторы, специфически соединяющиеся с определенным хемоэффектором, при этом молекула хемоэффектора не изменяется, а в молекуле рецептора происходят конформационные изменения. Рецепторы расположены неравномерно по всей поверхности клетки, а сконцентрированы на одном из полюсов. Состояние рецептора отражает внеклеточную концентрацию соответствующего эффектора.
Хемотаксис имеет приспособительное значение. Например, формы холерного вибриона с нарушенным хемотаксисом оказываются менее вирулентными.
Аэротаксис — нуждающиеся в молекулярном кислороде бактерии скапливаются вокруг пузырьков воздуха, попавших под покровное стекло.
Фототаксис— движение к свету или от него, свойствен фототрофным бактериям, использующим свет в качестве источника энергии.
Магнитотаксис — способность водных бактерий, содержащих кристаллики железосодержащих минералов, плыть вдоль линий магнитного поля Земли.
Термотаксис — движение в сторону изменения температуры, что имеет большое значение для некоторых патогенных бактерий.
Вискозитаксис— способность реагировать на изменение вязкости раствора. Обычно бактерии стремятся в среду с большей вязкостью, что имеет большое значение для патогенных видов.
Скольжение бактерий. Способность к скольжению с небольшой скоростью (2–11 мкм/с) по твердому или вязкому субстрату обнаружена у некоторых прокариот, напр., микоплазм.
Существуют несколько гипотез, объясняющих скользящее движение. Согласно гипотезе реактивного движения оно обусловлено выделением слизи через многочисленные слизевые поры в КС, в результате чего клетка отталкивается от субстрата в направлении, противоположном направлению выделения слизи. Согласно гипотезе «бегущей волны» скользящее движение у подвижных безжгутиковых форм связано с наличием между пептидогликановым слоем и наружной мембраной КС тонкого белкового слоя из упорядоченно расположенных фибрилл, аналогичных нитям жгутиков. Вращательное движение фибрилл, «запускаемое» этими структурами, приводит к появлению на поверхности клетки «бегущей волны» (движущихся микроскопических выпуклостей КС), в результате чего клетка отталкивается от субстрата. Наконец, у некоторых скользящих бактерий описаны структуры, напоминающие базальные тела жгутиковых форм.
Функции жгутиков:
1. Обеспечивают адгезию — начальную стадию инфекционного процесса.
2. Обеспечивают подвижность бактерий.
3. Определяют антигенную специфичность, это Н-антиген.
Выявление жгутиков:
1. Фазовоконтрастная микроскопия нативных препаратов («раздавленной» и «висячей» капли). Микроскопически подвижность определяют у клеток суточной культуры. Для того чтобы отличить подвижность от пассивного броуновского движения, к капле исследуемой культуры добавляют каплю 5 %–ного водного раствора фенола, активное движение в этом случае прекращается.
2. Темнопольная микроскопия нативных препаратов.
3. Световая микроскопия окрашенных красителями или металлами препаратов. Так как жгутики очень легко повреждаются при приготовлении препарата, в повседневной практике эти методы используется редко.
Для окраски жгутиков используют клетки, выращенные на скошенном агаре. Бактериальной петлей отбирают клетки, находящиеся у конденсационной воды и осторожно переносят в стерильную дистиллированную воду такой же температуры, что и температура инкубирования бактерий на скошенном агаре, а бактерии с петли не стряхивают, а осторожно погружают в воду. Пробирку с бактериями оставляют при комнатной температуре на 30 мин. Используют химически чистое (вымытое в хромовой смеси) стекло, на которое наносят 2–3 капли суспензии. Суспензию распределяют по поверхности стекла, осторожно его наклоняя. Высушивают препарат на воздухе.
Жгутики очень тонкие, поэтому их можно обнаружить только при специальной обработке. Вначале при помощи протравки достигается разбухание и увеличение их размера, а затем производится окраска препарата, благодаря чему они становятся видимыми при световой микроскопии.
Чаще используют метод серебрения по Морозову (рис. 10):
– препарат фиксируют раствором ледяной уксусной кислоты 1 минуту, промывают водой;
– наносят раствор таннина (дубящий, делающий жгутики более плотными) на 1 мин, промывают водой;
– обрабатывают препарат при подогревании импрегнирующим раствором азотнокислого серебра 1–2 мин, промывают водой, высушивают и микроскопируют.
При микроскопии видны темно-коричневые клетки и более светлые жгутики.
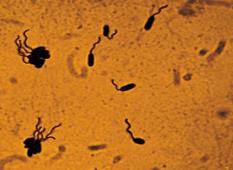
Рис. 10. Выявление жгутиков методом серебрения
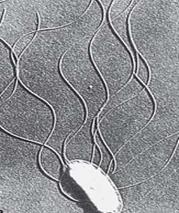
Рис. 11. Выявление жгутиков
методом электронной микроскопии
4. Электронная микроскопия препаратов, напыленных тяжелыми металлами (рис. 11).
5. Косвенно — по характеру роста бактерий при посеве в полужидкий 0,3 %–ный агар. После инкубирования посевов в термостате в течение 1–2 сут отмечают характер роста бактерий:
– у неподвижных бактерий (напр., S. saprophyticus) наблюдается рост по ходу укола — «гвоздь», а среда прозрачна;
– у подвижных бактерий (напр., Е. со1i)наблюдается рост в стороны от укола, по всему столбику агара — «елочка», и диффузное помутнение среды.
Дата добавления: 2015-04-25 ; Просмотров: 5540 ; Нарушение авторских прав? ;
Нам важно ваше мнение! Был ли полезен опубликованный материал? Да | Нет
| Состояние | отпатрулирована |

Жгу́тик (англ. Flagellum ) — поверхностная структура, присутствующая у многих прокариотических (бактерий и архей) и эукариотических клеток и служащая для их движения в жидкой среде или по поверхности твёрдых сред. Жгутики прокариот и эукариот принципиально различаются и по структуре, и по источнику энергии для движения и по механизму движения. Жгутики могут присутствовать на клетках по одному или во множестве, они имеются у многих протистов, зооспор и гамет. Эукариотический жгутик представляет собой вырост клетки, окружённый мембраной, с элементами цитоскелета внутри, который осуществляет биения за счёт энергии гидролиза АТФ. Бактериальный жгутик, как и жгутик архей, вмонтирован в оболочку клетки и вращается за счёт энергии трансмембранного градиента протонов или ионов натрия.
Помимо плавания, жгутики играют важную роль и в других процессах в жизни бактерий. Они участвуют в образовании биоплёнок, обеспечивают контакт клеток с субстратом, облегчают колонизацию хозяина симбиотическими бактериями [en] , служат одним из факторов вирулентности, запускают иммунный ответ организма хозяина.
Содержание
Жгутики эукариот [ править | править код ]

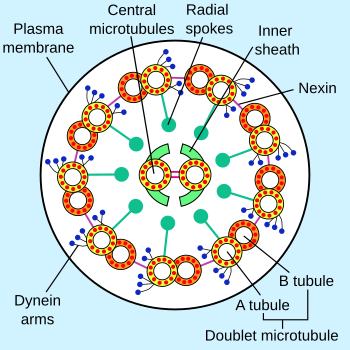
Жгутик эукариот представляет собой тонкий вырост на поверхности клетки, покрытый клеточной мембраной и содержащий аксонему — сложный структурный элемент, представляющий собой систему микротрубочек. Жгутиками снабжены гаметы у многих организмов, зооспоры и клетки многих протистов. Жгутик осуществляет движение по типу ундуляции [en] , совершая 10—40 оборотов в секунду. Энергия для движения эукариотического жгутика добывается при гидролизе АТФ. Внешнюю, внеклеточную часть жгутика называют ундулиподией [1] .
Ундулиподия обычно имеет одинаковую толщину по всей длине, которая составляет от 200 до 250 нм. Почти у всех свободноживущих протистов ундулиподии несут чешуйки или мастигонемы [en] (нитевидные структуры, отходящие от поверхности жгутика) [2] . Структурная основа ундулиподии представлена аксонемой, которая состоит из 9 пар периферических микротрубочек (дублетов) и двух центральных микротрубочек. Эта формула (9 × 2 + 2) характерна для всех эукариот, хотя известны и исключения (чаще всего редуцируется центральная пара микротрубочек, но иногда в аксонеме появляются дополнительные микротрубочки). Компоненты аксонемы имеют сложный состав и образованы более чем 250 белками. Две центральные микротрубочки имеют диаметр 25 нм и располагаются на расстоянии 30 нм друг от друга [1] , они одеты общим чехлом из тонкого материала. При помощи радиальных спиц чехол соединяется с периферическими дублетами микротрубочек. Каждый дуплет образован микротрубочками A и B, причём радиальные спицы крепятся к A-микротрубочкам. От A-микротрубочек также отходят динеиновые мостики и нексиновые [en] ручки. Динеиновые ручки направлены к соседнему дублету и обусловливают изгибание аксонемы, обеспечивая скольжение дублетов относительно друг друга. Нексиновые мостики нужны для прочной связки соседних дублетов в переходной зоне аксонемы, которая не подвергается изгибаниям. Иногда жгутики эукариот несут дополнительные структурные элементы, например, параксиальный тяж, имеющийся у некоторых протистов (динофлагеллят, эвгленовых и других). Параксиальный тяж состоит из микрофиламентов и идёт вдоль аксонемы [3] .
На уровне выхода жгутика из клетки находится так называемая переходная зона аксонемы. Аксонема крепится к кинетосоме, или базальному телу, располагающемуся под клеточной мембраной. Кинетосома представляет собой полый цилиндр, стенка которого состоит из 9 триплетов микротрубочек, соединённых, как правило, фибриллярными мостиками. В состав триплета, помимо аксонемных A- и B-микротрубочек, входит дополнительная C-микротрубочка, прилегающая к B-микротрубочке. В центре проксимальной части кинетосомы находится ось со спицами, которая также имеется у центриолей [4] .
От кинетосом непосредственно отходят так называемые корешки — фибриллярные или микротрубочковые структуры. Иногда они соединяются с кинетосомами короткими фибриллярными связками или начинаются в слое аморфного материала, окружающего кинетосому. Корешки обоих типов служат для закрепления жгутика в теле клетки [5] .
Как правило, эукариотические клетки несут от 1 до 8 жгутиков, хотя среди протистов известно немало многожгутиковых форм. Жгутиконосных протистов подразделяют на четыре основные группы: изоконты несут одинаковые жгутики с одинаковым способом биения; анизоконты обладают жгутиками разной длины, но они также имеют одинаковый вид и способ биения; у гетероконтов имеется два разных по внешнему виду и расположению жгутика с разными способами биения; наконец, стефаноконтные формы имеют венчик из 30—40 жгутиков на переднем конце клетки [6] .
Жгутики бактерий [ править | править код ]

Жгутиками обладают до 80 % известных видов бактерий [7] . Количество и расположение жгутиков различно у разных видов. У некоторых бактерий есть всего один жгутик (монотрихи), у других два жгутика располагаются на двух противоположных концах клетки (амфитрихи), у третьих на полюсах клетки находятся пучки жгутиков (лофотрихи), а у четвёртых жгутики покрывают всю поверхность клетки (перитрихи). У спирохет жгутик находится в периплазматическом пространстве между двумя мембранами. Клетки спирохет имеют характерную извитую форму, которая меняется при движении [8] .

Строение жгутика бактерий фундаментально отличается от строения жгутика эукариот. Если эукариотический жгутик представляет собой вырост клетки, окружённый мембраной, с элементами цитоскелета внутри, который движется за счёт энергии гидролиза АТФ, то бактериальный жгутик (как и жгутик архей) вмонтирован в оболочку клетки и вращается за счёт энергии трансмембранного градиента протонов или ионов натрия. Кроме того, в отличие от эукариотических жгутиков, основной тип движения которых — биение, жгутики бактерий вращаются [9] . Жгутики бактерий и архей состоят из двух независимых частей — ротора и статора [10] .
Жгутик вращается попеременно по часовой стрелке и против неё. Частота вращения постоянна для конкретной клетки и составляет от 250 до 1700 Гц (то есть от 1,5⋅10 4 до 10 5 оборотов в минуту) [10] .
Строение жгутика лучше всего изучено у кишечной палочки Escherichia coli и Salmonella enterica серотип Typhimurium [en] . Гены, ответственные за движение у этих бактерий, подразделяют на три группы:
- fla-гены (от англ. flagellum ) кодируют субъединицы жгутика и компоненты, необходимые для его биогенеза. В эту группу входит около 50 генов;
- mot-гены (от англ. motility — подвижность) отвечают за вращение мотора жгутика;
- che-гены (от англ. chemotaxis — хемотаксис) обеспечивают плавание как адаптивную реакцию [11] .

К числу основных структурных компонентов бактериального жгутика относят:
- филамент (нить) — жёсткая нитевидная структура длиной до 15 мкм и диаметром около 20 нм, располагающаяся за пределами цитоплазматического компартмента;
- базальное тело — опорная структура, которая у грамотрицательных бактерий представлена муреиновым чехлом совместно с внешней мембраной, а у грамположительных она представлена только муреиновым чехлом. В состав базального тела входит система секреции III типа, которая служит для экспорта из клетки субъединиц жгутика, а также мотор жгутика.
- крюк длиной около 55 нм и диаметром около 30 нм, который представляет собой гибкое сочленение между филаментом и базальным телом [12] .
Рабочей частью жгутика является филамент, или нить. Филамент представляет собой спирализованный цилиндр с внутренним каналом диаметром около 3 нм. Филамент состоит из около 20 тысяч идентичных субъединиц белка флагеллина (FliC). У некоторых бактерий, например, Helicobacter pylori, Caulobacter crescentus [en] , Sinorhizobium meliloti [en] и Campylobacter coli [en] в состав филамента входят флагеллиновые субъединицы двух типов. У ряда бактерий флагеллиновые субъединицы гликозилированы. Флагеллиновые субъединицы в составе филамента уложены спирально, на один оборот спирали приходится 11 субъединиц, поэтому они образуют 11 параллельных протофиламентов, проходящих под небольшим углом к оси филамента. Форма филамента определяется аминокислотной последовательностью входящих в его состав флагеллинов, ионной силой и pH раствора, в котором перемещается клетка. Когда мотор вращается против часовой стрелки, филамент принимает форму левозакрученной спирали, а когда по часовой — правозакрученной. На дистальном конце филамента находится шапочка (кэп), который имеет форму диска и состоит из пяти субъединиц белка Fl >[13] .
Филамент соединяется с базальным телом посредством крюка, который образован субъединицами FlgE, вспомогательными белками FlgKL и кэпирующим белком FlgD, который удаляется при завершении сборки крюка. Крюк играет роль гибкого сочленения, благодаря которому жёсткая нить может менять своё положение по отношению к поверхности клетки [14] .
Базальное тело встроено в клеточную стенку и у грамотрицательных бактерий состоит из следующих частей:
- оси (белки FlgBCFG и FliE), которая дистальной частью связана с крюком и нитью, а проксимальной частью — с MS-кольцом;
- L-кольца (белки FlgH), которое вставлено во внешнюю мембрану и функционирует как поддерживающая втулка для вращающейся оси;
- P-кольца (белки FlgI), интегрированного в муреиновый чехол и выполняющего роль поддерживающей втулки для оси;
- MS-кольца (26 субъединиц FliF), к которому причленяются проксимальные субъединицы оси (FliE), субъединицы ротора (FliG), переключатель направления вращения (состоит из 37 субъединиц FliM и 110 субъединиц FliN);
- ротора (44 субъединицы FliG), который залегает в основании MS-кольца;
- C-кольца (белки FliMN), связанного с ротором и задействованного в смене направления движения [15] .
Так как биосинтез субъединиц жгутика требует много клеточных ресурсов, он регулируется специальным каскадом, в который у E. coli и Salmonella enterica серовар Typhimurium входят три класса генов. Аппарат для экспорта субъединиц жгутика состоит из субъединиц FlhAB, FliOPQR и FliHIJ, выступает в цитоплазму, проходит через центральное отверстие ротора и частично погружён в MS-кольцо [16] .
Характер плавания бактериальной клетки зависит от количества и расположения жгутиков. У монотрихов, например, Pseudomonas aeruginosa, жгутик, вращающийся по часовой стрелке, толкает клетку вперёд, а против — тянет её за собой. У некоторых бактерий жгутик вращается только по часовой стрелке. У бактерий со множеством жгутиков (политрихов) при плавании жгутики сплетаются в жгут, вращающийся против часовой стрелки. При вращении по часовой стрелке жгут расплетается, и из-за независимого вращения отдельных жгутиков клетка начинает «кувыркаться» и непредсказуемо меняет направление движения [17] .
Помимо плавания, жгутики играют важную роль и в других процессах в жизни бактерий. Они участвуют в образовании биоплёнок, обеспечивают контакт клеток с субстратом, облегчают колонизацию хозяина симбиотическими бактериями [en] , а для патогенных бактерий жгутики служат одним из факторов вирулентности, так как облегчают проникновение в тело хозяина, перемещение в вязкой межклеточной среде, проникновение в щели между клетками эндотелия. Жгутики бактерий высоко иммуногенны и запускают иммунный ответ организма хозяина, поэтому компоненты жгутика используют для пассивной иммунизации [18] .
У некоторых бактерий рода Vibrio (в частности, Vibrio parahaemolyticus [en] [19] ) и некоторых протеобактерий, таких как Aeromonas, имеются две различные жгутиковые системы, белковые компоненты которых кодируются различными наборами генов, а для вращения используются разные ионные градиенты. Полярные жгутики, относящиеся к первой жгутиковой системе, присутствуют постоянно и обеспечивают подвижность в потоке жидкости, а боковые жгутики, относящиеся ко второй жгутиковой системе, экспрессируются только тогда, когда сопротивление окружающей жидкости так велико, что полярные жгутики не могут вращаться. Благодаря этому бактерии могут скользить по различным поверхностям и в вязкой жидкости [20] [21] [22] [23] [24] [25] .
Жгутики архей [ править | править код ]

Жгутик архей (археллум) служит, как и у других организмов, преимущественно для перемещения в жидкой среде. Структурно археллум схож с пилями IV типа [27] . Большинство генов, белковые продукты которых образуют археллум, образуют единый оперон fla. Этот оперон содержит от 7 до 13 генов, продукты которых участвуют в сборке и работе археллума [28] . Структурные компоненты археллума называют археллинами, эти белки кодируются генами flaA и flaB; моторные компоненты кодируются генами flaI, flaJ и flaH. В этом опероне также закодированы вспомогательные белки FlaG, FlaF и FlaX, а также сигнальные белки FlaC, FlaD и FlaE. Генетический анализ различных видов архей показал, что все перечисленные белки необходимы для сборки археллума [29] [30] [31] [32] [33] . У эвриархеот имеется почти полный набор генов fla, а у представителей типа кренархеот один или два из них могут отсутствовать. Для созревания археллинов необходима пептидаза препилин [en] (у эвриархеот она известна как PibD, у кренархеот — FlaK), однако она не входит в состав fla-оперона [34] .
Хотя археллум во многих отношениях значительно отличается от жгутика бактерий, основная его функция та же — перемещение клетки в жидкой среде [33] [35] [36] или на влажных поверхностях [37] [38] . Подобно бактериальному жгутику [39] [40] , археллум участвует в прикреплении клеток к субстрату и межклеточной коммуникации [41] [42] , однако, в отличие от жгутиков бактерий, не задействован в образовании биоплёнок [43] .
Схема строения жгутика бактерий.
Жгутики бактерий определяют подвижность бактериальной клетки. Жгутики представляют собой тонкие нити, берущие начало от ЦПМ, имеющие большую длину, чем сама клетка. Толщины жгутиков 12-20 нм, длина 3-12 мкм.
Жгутики состоят из сократительного белка типа миозина – флагеллина (от. лат. flagellum – жгутик), обладающего антигенной специфичностью. Субъединицы флагеллина закручены в виде спирали.


Жгутик состоит из трех компонентов – спиральной жгутиковой нити (филамента) постоянной толщины, крючка (колена) и базального тельца. Крючок, к которому присоединена жгутиковая нить, имеет длину 30-45 нм и состоит из отличающегося от флагеллина белка. Он соединен с базальным тельцем, которое располагается в оболочке (в клеточной стенке и ЦПМ). Нити жгутиков приводит в движение мембранный шарнироподобный базальный крюк. Базальное тельце состоит из центрального стержня, заключенного в систему особых колец. Кольца выполняют роль «приводного диска» и «подшипника» на внутренней поверхности пептидогликанового слоя. Вся конструкция выполняет функцию хемомеханического преобразователя (флагеллиновый мотор).
У грамотрицательных бактерий две пары колец: внешняя (кольца L и Р) и внутренняя (кольца S и М). Кольца L и Р расположены внутри клеточной стенки (кольцо L в ЛПС, а кольцо Р – в слое пептидогликана). Они выполняют, очевидно, роль втулки для стержня. Внутренняя пара (кольца S и М) фиксирована на ЦПМ, причем кольцо S располагается в периплазматическом пространстве, а кольцо М – на ЦПМ или в ней. Жгутики грамположительных бактерий, имеющих более толстую гомогенную клеточную стенку, содержат только одну пару колец – S и М.
Вращение жгутика в клеточной стенке происходит из-за вращательного движения колец S и М относительно друг друга и обеспечивается за счет энергии трансмембранного градиента ионов водорода или натрия. Благодаря такому вращению происходит движение бактерий в наиболее благоприятном для них направлении. Жгутиковый аппарат обладает особым бинарным переключателем, который позволяет менять направление вращения жгутиков против часовой стрелки на противоположное. Таким образом, бактерии, получив химический сигнал из окружающей среды, изменяют направление движения и выбирают оптимальные условия обитания.
Модель движения жгутика.

Число жгутиков у бактерий различных видов варьирует от одного (монотрих) у холерного вибриона до десятка и сотен жгутиков, отходящих по периметру бактерии (перитрих) у кишечной палочки, протея и др. Лофотрихи имеют пучок жгутиков на одном конце клетки. Амфитрихи имеют по одному жгутику или пучку жгутиков на противоположных концах клетки.
Расположение жгутиков у бактерий.

Жгутики выявляют при помощи электронной микроскопии препаратов, напыленных тяжелыми металлами, или в световом микроскопе после обработки специальными методами, основанными на протравливании и адсорбции различных веществ, приводящих к увеличению толщины жгутиков (например, после серебрения). Подвижность бактерий определяют при помощи фазово-контрастной или световой микроскопии «раздавленной» или «висячей» капли).

Vibrio cholerae. Фрагменты клеток с полярно расположенным жгутиком (Ж). Электронная микроскопия, х130000.